Модельные объекты
Раздел о тех видах живых организмов, которые используются в науке как модельные объекты в исследовании биологических процессов.
Арабидопсис
 Arabidopsis Thaliana
Arabidopsis Thaliana
Arabidopsis Thaliana - вид семейства Brassicaceae, родственник горчицы и капусты. Однако, в отличие от них является сорным растением. Благодаря своим особенностям является удобным объектом исследований:
• Развитие, размножение, заболевание и реакции на воздействие окружающей среды такие же как и у съедобных цветковых растений.
• Малый размер растений (30-40 см), быстрое воспроизводство (5-8 недель от проростания до созревания семян) и образование тысяч семян от одного растения упрощают выращивание в лабораторных условиях.
• Малый размер генома в сравнении с другими растениями (125 млн пн), малое число хромосом (5 пар) и малое число повторенных последовательностей. Обнаружено 25500 генов.
• Самоопыляемость растения позволяет выращивать гомозиготные линии.
• Трансформация при помощи бактерии Agrobacterium tumefaciens, привела к накоплению линий с различными мутациями.
Примерно 115Mb из 125 Mb секвенировано
Сначала растения формируют розетку лежащую на земле затем появляются цветы на стебле образуя концевую кисть.
Листья небольшие, примерно, 1-5 см в длину, слегка зубчатые, со звездчатым пушком. Семена цилиндрические 1мм в ширину и 1-2 см в длину находятся в стручке.
Бактерии
Вироиды
Вирусы
Наследственный материал вирусов.
| одноцепочечные | двухцепочечные | ||
| + | - | ||
| РНК | ретровирусы ВИЧ |
грипп бешенство корь |
|
| ДНК | герпес оспа гепатит B |
||
Ретроврусы.
d~100нм
РНК ретровирусов состоит из следующих элементов: str5-U5-gag-pol-env-onc-str3-U3-polyA
gag - белок нуклеоида
pol - обратная транскриптаза
env - белок капсида
onc - онкоген-злокачественная трансформация клеток
str5 = str3 - short terminal repeat - короткий концевой повтор,U3,U5-уникальные послед-ти (U5-80н,U3-200н),PB-primer binding site - участок связывания затравки схема обратной транскрипции ретровирусов
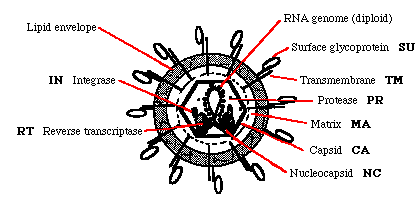
рис.1 Строение ретровируса
MA (Matrix) - белок матрикса (gag); lines envelope
CA (Capsid) - белок капсида (gag ген); защищает кор; 1шт.
NC (Nucleocapsid) - белок капсида(gag gene); защищает геном; формирует кор.
PR (Protease) - необходима для расщепления gag белка во время созревания.
RT (Reverse transcriptase) - Reverse transcribes the RNA genome; also has RNAseH activity.
IN (Integrase) - Encoded by the pol gene; needed for integration of the provirus.
SU (Surface glycoprotein) - наружная поверхность гликопротеинов; главный вирусный антиген.
TM (Transmemb protein) - внутренний компонент гликопротеиновой оболочки.

рис.2 Схема обратной транскрипции ретровирусов - образование ДНК по матрице одноцепочечной РНК.
Дрожжи
Saccharomyces cerevisiae - почкующиеся дрожжи (budding yeast). Клетки округлой формы, 5-10 мкм в диаметре.
Schizosaccharomyces pombe - делящиеся дрожжи (fission yeast). Клетки палочковидной формы, 2-3x7-12 мкм.
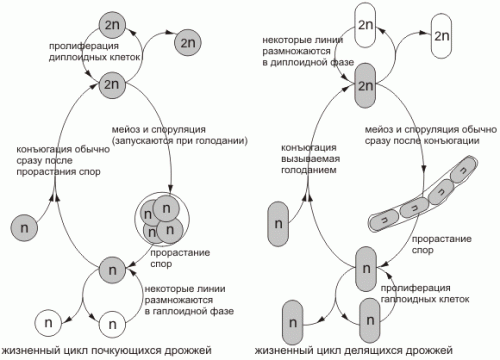 Жизненные циклы дрожжей
Жизненные циклы дрожжей
рис.1 Жизненные циклы почкующихся дрожжей (Saccharomyces cerevisiae) и делящихся дрожжей (Schizosaccharomyces pombe). Доли жизненного цикла, проводимые в гаплоидной и в диплоидной фазе, меняются от вида к виду и в зависимости от условий среды. При обилии пищи нормальные разновидности дикого типа почкующихся дрожжей размножаются как диплоидные клетки с продолжительностью жизненного цикла около двух часов. При голодании же они претерпевают мейоз с образованием гаплоидных спор, которые в благоприятных условиях прорастают, превращаясь в гаплоидные клетки. Эти клетки в зависимости от условий среды и генотипа либо делятся, либо сливаются (конъюгируют) в фазе G1, вновь образуя диплоидные клетки. Наоборот, делящиеся дрожжи обычно размножаются в гаплоидном состоянии; при недостатке пищи гаплоидные клетки сливаются с образованием диплоидных клеток, которые быстро проходят мейоз и споруляцию с восстановлением гаплоидной фазы. Наиболее широко используемые лабораторные штаммы почкующихся дрожжей - это мутанты, которые, подобно делящимся дрожжам, размножаются главным образом в гаплоидной фазе. (Альбертс, 1994)
Дрозофила
Дрозофила - род содержащий более 500 видов, некоторые из которых приведены здесь
Благодаря своим особенностям дрозофила является удобным объектом генетических сследований:
1. Короткое время размножения (десять дней от яйца до половозрелой мухи)
2. Большое число потомков
3. Малый размер и неприхотливость в содержании
4. Большое количество спонтанных мутаций (см. обзор Мутации дрозофилы)
5. Наличие политенных хромосом (см. обзор Политенные хромосомы)
ЖИЗНЕННЫЙ ЦИКЛ ДРОЗОФИЛ.
Жизненный цикл дрозофилы при 25 °C занимает 10 дней, при 18 °C — месяц.
Самки откладывают около 400 яиц, каждое из которых порядка 0,5 мм в длину. Яйца раскрываются через 24 часа. Вылупившиеся личинки растут на протяжении 5 дней, дважды линяя за это время: через 24 и 48 часов после рождения. Затем личинки выползают на поверхность и подсыхая покрываются твердой оболочкой - пупарий и претерпевают пятидневную стадию метаморфоза, в результате которого возникают взрослые особи – имаго.
Самки могут быть оплодотворены только один раз за свою жизнь, после чего они непрерывно откладывают яйца, при этом сперма хранится внутри тела самки. После нескольких часов после вылета (5-8) самки остаются стерильными. Именно в этот промежуток экспериментатор собирает вылетевших мух для скрещиваний.
Глаз дрозофилы состоит из 800 омматидиев, каждый из которых состоит из 8 проторецепторных клеток, поддерживающих клеток, пигментных клеток и роговицы. Мухи дикого типа имеют красный цвет глаз. Каждая фоторецепторная клетка состоит из клеточного тела, содержащего ядро и рабдомера состоящего из большого количества мембран содержащих родопсин. Фоторецепторные клетки экспрессируют различные изоформы родопсина. R1-R6 фоторецепторные клетки экспрессируют Rhodopsin1 (Rh1), поглощающий голубой цвет (480 nm). R7 и R8 клетки экспрессируют Rh3 или Rh4, поглащающие ультрафиолет (345 nm и 375 nm), и Rh5 или Rh6,поглощающие голубой (437 nm) и зеленый (508 nm) свет.
Пигментация глаз определяется наличием двух генетически независимых пигментных систем, красной и коричневой. Мухи дикого типа имеют кирпично-красный цвет глаз, соответствующий присутствию обеих систем. Коричневая составляющая образуется из двух пигментов омматина и оммина. Красная составляющая образуется из множества пигментов, называемых птеринами. При мутации затрагивающей обе пигментные системы цвет глаз становится белым. Например мутация white, при которой нарушается транспортировка любых пигментов внутри клеток глаза.
ОТЛИЧИЕ САМЦОВ И САМОК.
ПОВЕДЕНИЕ ДРОЗОФИЛ.
 Спаривающиеся дрозофилы
Спаривающиеся дрозофилы
Мутации дрозофилы
Мутации дрозофилы (Drosophila melanogaster).
 Bar (B) - доминантная мутация
Bar (B) - доминантная мутация
 white (w) - рецессивная мутация гена white в Х-хромосоме. Ген переноса пигментов в клетках глаза не работает совсем, глаза имеют белый цвет.
white (w) - рецессивная мутация гена white в Х-хромосоме. Ген переноса пигментов в клетках глаза не работает совсем, глаза имеют белый цвет.
 white apricot (wa) -
white apricot (wa) -
 yellow (y) - Желтое тело и крылья. Мутация гена yellow в X-хромосоме.
yellow (y) - Желтое тело и крылья. Мутация гена yellow в X-хромосоме.
 bitorax (bx)
bitorax (bx)
 Cut
Cut
 lz
lz
 sc
sc
 ebony (e) - Черное тело. Мутация гена ebony в третьей хромосоме.
ebony (e) - Черное тело. Мутация гена ebony в третьей хромосоме.
scarlett (sc) - Алые глаза.
 vg
vg
short-wingled - Рецессивная мутация в гене (?) во второй хромосоме. Крылья короткие, мухи с такой мутацией не способны летать.мутация гена Curly во второй хромосоме. Крылья загнуты кверху.
black (b) - мутация второй хромосомы
forked (f) - мутация Х-хромосомы. Вильчатые крылья
orange-eyed - мутация гена white. Глаза оранжевого цвета. Ген white работает лишь частично выполяя свою функцию
eyeless (ey) - мутация гена в 4 хромосоме, регулирующем построение глаза на личиночной стадии. Отсутствие глаз.
polished (lac) - мутация в 4 хромосоме.Меняется глаз
антенапедия, направляющего некоторые клетки тела формировать ноги. Ноги образуются на голове вместо пары антенн.
brown (bw) - Коричневый цвет глаз. Мутация второй хромосомы
ruby (rb) - Цвет глаз. Х-хромосома
sepia (se) - глаз, 3
vermillion (v)- Х-хромосома, глаз
stubble (Sb) - короткие щетинки
tabby (Tb)
apterous - совсем нет крыльев
Список всех видов Дрозофилы
Drosophila abjuncta
Drosophila acrostichalis
Drosophila acutilabella
Drosophila adiastola
Drosophila adventitia
Drosophila agitona
Drosophila aglaia
Drosophila alabamensis
Drosophila albifacies
Drosophila aldrichi
Drosophila algonquin
Drosophila amydrospilota
Drosophila ananassae
Drosophila ancyla
Drosophila anomalipes
Drosophila anthrax
Drosophila apicipuncta
Drosophila apicisetae
Drosophila apodasta
Drosophila apoxyloma
Drosophila aquila
Drosophila araiotrichia
Drosophila artigena
Drosophila asketostoma
Drosophila attenuata
Drosophila azteca
Drosophila badia
Drosophila baja
Drosophila balioptera
Drosophila basdeni
Drosophila basimacula
Drosophila basisetosa
Drosophila beardsleyi
Drosophila bicondyla
Drosophila bifurca
Drosophila bimaculata
Drosophila bipolita
Drosophila biseriata
Drosophila bizonata
Drosophila boletina
Drosophila borealis
Drosophila bostrycha
Drosophila brevicarinata
Drosophila brevicilia
Drosophila brevissima
Drosophila brevitarsus
Drosophila bridwellius
Drosophila bromeliae
Drosophila brooksi
Drosophila brunneifrons
Drosophila brunneisetae
Drosophila busckii
Drosophila caccabata
Drosophila californica
Drosophila canapalpa
Drosophila canipolita
Drosophila canuta
Drosophila capitata
Drosophila carbonaria
Drosophila cardini
Drosophila carnosa
Drosophila carsoni
Drosophila casteeli
Drosophila chaetopeza
Drosophila chagrinensis
Drosophila ciliaticrus
Drosophila cilifemorata
Drosophila cinerea
Drosophila clavata
Drosophila clavitibia
Drosophila clydonia
Drosophila cnecopleura
Drosophila colorata
Drosophila comatifemora
Drosophila conformis
Drosophila confutata
Drosophila conjectura
Drosophila conspicua
Drosophila contorta
Drosophila cracens
Drosophila crassifemur
Drosophila crucigeraur
Drosophila curticilia
Drosophila curvitibia
Drosophila dasycnemia
Drosophila deflecta
Drosophila deltaneuron
Drosophila demipolita
Drosophila denotata
Drosophila desertorum
Drosophila diffusa
Drosophila diminuens
Drosophila disjuncta
Drosophila dissita
Drosophila disticha
Drosophila distinguenda
Drosophila dobzhanskii
Drosophila dolomata
Drosophila dorsigera
Drosophila dorsociliata
Drosophila dracaenae
Drosophila dumalis
Drosophila duncani
Drosophila elliptica
Drosophila engyochracea
Drosophila enoplotarsus
Drosophila eremophila
Drosophila eumecothrix
Drosophila euronotus
Drosophila eurypeza
Drosophila expansa
Drosophila falleni
Drosophila fasciculisetae
Drosophila fastigata
Drosophila felix
Drosophila flavibasis
Drosophila flaviceps
Drosophila flavisternum
Drosophila flavomontana
Drosophila flavopinicola
Drosophila flexa
Drosophila floricola
Drosophila fragilis
Drosophila freycinetiae
Drosophila fulvalineata
Drosophila fundita
Drosophila funebris
Drosophila fungicola
Drosophila furva
Drosophila furvifacies
Drosophila fuscifrons
Drosophila fuscoamoeba
Drosophila fuscoapex
Drosophila fusticula
Drosophila gilvilateralis
Drosophila grimshawi
Drosophila grisea
Drosophila guttifera
Drosophila haleakalae
Drosophila hamatofila
Drosophila hawaiiensis
Drosophila hexachaetae
Drosophila hexastigma
Drosophila hirticoxa
Drosophila hirtitarsus
Drosophila hirtitbia
Drosophila humeralis
Drosophila hydei
Drosophila hydeoides
Drosophila iki
Drosophila illusiopolita
Drosophila imitator
Drosophila immigrans
Drosophila imparisetae
Drosophila improcera
Drosophila incognita
Drosophila incompleta
Drosophila inedita
Drosophila infuscata
Drosophila innubila
Drosophila insignita
Drosophila intermedia
Drosophila involuta
Drosophila iroquois
Drosophila ischnotrix
Drosophila joycei
Drosophila kauluai
Drosophila kikkawai
Drosophila krassi
Drosophila lacicola
Drosophila laciniosa
Drosophila lanaiensis
Drosophila larifuga
Drosophila latifasciaeformis
Drosophila latigena
Drosophila lemniscata
Drosophila leonis
Drosophila limpiensis
Drosophila lonchoptera
Drosophila longala
Drosophila longicornis
Drosophila lowei
Drosophila luteola
Drosophila lutzii
Drosophila macrochaetae
Drosophila macropolia
Drosophila macroptera
Drosophila macrospina
Drosophila magnabadia
Drosophila magnafumosa
Drosophila magnaquinaria
Drosophila magnimacula
Drosophila magnipalpa
Drosophila mahican
Drosophila mainlandi
Drosophila mathisi
Drosophila mecocnemia
Drosophila mediana
Drosophila megasticta
Drosophila melanderi
Drosophila melanica
Drosophila melanissima
Drosophila melanogaster (fruit fly)
Drosophila melanoloma
Drosophila melanopedis
Drosophila melanosoma
Drosophila melanura
Drosophila mercatorum
Drosophila meridiana
Drosophila mettleri
Drosophila micromelanica
Drosophila mimica
Drosophila mimiconformis
Drosophila mimiconfutata
Drosophila miranda
Drosophila mitchelli
Drosophila mojavensis
Drosophila molokaiensis
Drosophila montana
Drosophila mulleri
Drosophila munda
Drosophila musae
Drosophila musaphilia
Drosophila mycetophila
Drosophila nanella
Drosophila narragansett
Drosophila nasalis
Drosophila nebulosa
Drosophila neglecta
Drosophila neorepleta
Drosophila neutralis
Drosophila nigella
Drosophila nigra
Drosophila nigripalpus
Drosophila nigritarsus
Drosophila nigrocirrus
Drosophila nigrohalterata
Drosophila nigrohydei
Drosophila nigromelanica
Drosophila nigropolita
Drosophila nigrospiracula
Drosophila novitskii
Drosophila occidentalis
Drosophila ochracea
Drosophila ohiensis
Drosophila olaae
Drosophila orbospiracula
Drosophila ordinaria
Drosophila oreas
Drosophila orestes
Drosophila pachea
Drosophila pachuca
Drosophila palustris
Drosophila parva
Drosophila paucipuncta
Drosophila paucula
Drosophila pectinitarsus
Drosophila pegasa
Drosophila peniculipedis
Drosophila peninsularis
Drosophila percnosoma
Drosophila perissopoda
Drosophila persimilis
Drosophila petalopeza
Drosophila picticornis
Drosophila pictiventris
Drosophila pilimana
Drosophila pinicola
Drosophila plumosa
Drosophila polita
Drosophila polliciforma
Drosophila pollinospadix
Drosophila polychaeta
Drosophila ponderosa
Drosophila populi
Drosophila praesutilis
Drosophila pretiosa
Drosophila proceriseta
Drosophila prodita
Drosophila projectans
Drosophila prolaticilia
Drosophila prolixa
Drosophila prominens
Drosophila propachuca
Drosophila propiofacies
Drosophila pseudoobscura
Drosophila punalau
Drosophila putrida
Drosophila pychnochaetae
Drosophila quadrisetae
Drosophila quasianomalipes
Drosophila quasiexpansa
Drosophila quinaria
Drosophila racemova
Drosophila recens
Drosophila reducta
Drosophila redunca
Drosophila rellima
Drosophila repleta
Drosophila residua
Drosophila retrusa
Drosophila rioensis
Drosophila ritae
Drosophila robusta
Drosophila rubidifrons
Drosophila rubrifrons
Drosophila ruburra
Drosophila rustica
Drosophila sabroskyi
Drosophila sadleria
Drosophila scolostoma
Drosophila seclusa
Drosophila semifuscata
Drosophila seminole
Drosophila seorsa
Drosophila septuosa
Drosophila setapex
Drosophila setiger
Drosophila setipalpus
Drosophila sharpi
Drosophila sigmoides
Drosophila simulans
Drosophila sordidapex
Drosophila spectabilis
Drosophila spenceri
Drosophila spicula
Drosophila stalkeri
Drosophila stenoptera
Drosophila subfunebris
Drosophila suboccidentalis
Drosophila subpalustris
Drosophila subsigmoides
Drosophila subviridis
Drosophila sucinea
Drosophila suffusca
Drosophila swezeyi
Drosophila taeniata
Drosophila tamashiroi
Drosophila taractica
Drosophila tarphytrichia
Drosophila tendomentum
Drosophila tenebrosa
Drosophila tenuipes
Drosophila testacea
Drosophila tetraspilota
Drosophila texana
Drosophila thoracis
Drosophila tira
Drosophila torula
Drosophila totonigra
Drosophila tranquilla
Drosophila transfuga
Drosophila trichaetosa
Drosophila tripunctata
Drosophila trispina
Drosophila truncipenna
Drosophila undulata
Drosophila unicula
Drosophila unipucntata
Drosophila varga
Drosophila variabilis
Drosophila varipennis
Drosophila velata
Drosophila velutinifrons
Drosophila venusta
Drosophila vicaria
Drosophila victoria
Drosophila villitibia
Drosophila villosipedis
Drosophila vinnula
Drosophila virilis
Drosophila wheeleri
Drosophila williamsi
Drosophila willistoni
Drosophila xanthognoma
Drosophila xanthosoma
Drosophila xuthoptera
Drosophila z-notata
По данным сайта: http://animaldiversity.ummz.umich.edu/site/index.html
- По данным сайта: http://animaldiversity.ummz.umich.edu/site/index.html
Крыса
Крысы (Rattus), род грызунов (семейство мышиных – Muridae), многочисленные виды которого распространены по всему земному шару. Серая крыса - пасюк (Rattus norvegicus) - дикая крыса, встречающийся в большинстве городов умеренной зоны, особенно в Европе, на севере США и в Канаде. Белых крыс (альбиносная форма рыжей) разводят в лабораториях и держат как домашних животных. Длина тела у разных видов составляет от 13 до 30 см, не считая чешуйчатого хвоста примерно такой же или большей длины. Масса пасюка средних размеров – примерно 0,7 кг, длина тела без хвоста – 23 см.
Резцы у крыс не имеют корней и постоянно растут. Их передняя сторона покрыта эмалью, задняя — менее прочная и стирается быстрее, чем передняя. Из-за неравномерного стачивания вершины резцов всегда острые. Крысы могут перегрызать такие твердые предметы, как бетон.
Крысы размножаются почти круглый год, и их численность может очень быстро расти. Половой зрелости самки достигают примерно в 3-месячном возрасте. Но до 6 месяцев, когда все были уже явно половозрелыми, к размножению приступает только около 1% самок. В следующие 6 месяцев начинает размножаться еще 7% самок. А 92% самок до годовалого возраста остаются яловыми. Чем старше становятся самки, тем выше их плодовитость — число детенышей в одном приплоде и число приплодов в году. Беременность длится обычно 21—22 день, число детенышей в помете от 3 до 12, в среднем 8. Крысята рождаются голыми и слепыми, но быстро развиваются. Глаза открываются у них примерно на 15-й день, а через 3 недели они самостоятельно ищут корм. В возрасте двух лет крыса перестает размножаться, а к трем годам дряхлеет и теряет зубы. В неволе крысы доживают до 3-5 лет.
Крысы – общественные животные, но между ними нередко вспыхивают драки. Самая уязвимая часть их тела – хвост; на нем часто видны шрамы от ран, нанесенных сородичами.
Дикиее крысы могут распространять сыпной тиф, бубонную чуму, желтуху, туляремию, бешенство и трихинеллез. Они особенно обильны на мусорных свалках и в канализационных системах, а инфекции переносят, загрязняя хранящиеся пищевые продукты.
Поведение диких крыс
Социальные отношения и система спаривания
Дикие крысы живут в колониях. Самки образуют небольшие группы (до шести животных). Одна группа имеет одну собственную систему нор (подробнее о норах ниже). У каждой самки есть отдельная гнездовая камера, и они могут растить своих крысят одиночно или заботиться об общих детенышах всей группой. Вскоре после перехода на самостоятельное питание молодые самцы уходят из колонии.
Система отношений самцов и система спаривания крыс изменяется в зависимости от плотности населения колонии. При низкой плотности крыса-самец монополизирует нору самок. Он защищает территорию от других самцов и спаривается только с самками своей группы. Такая форма половых отношений, когда один самец спаривается с несколькими представителями противоположного пола, носит название полигиния (разновидность полигамии).
При высокой плотности населения (более одной крысы на пяти кв. метрах) самец-хозяин уже не в состоянии защитить собственную
территорию от других самцов (потому что их становится слишком много). Это вызывает появление системы иерархии, т.е. один самец становится социально доминирующим, в то время как другие становятся социально зависимыми. Самцы больше не защищают свои норы и почти не враждуют между собой из-за самок. Вместо этого, когда у самки появляется течка, с ней спариваются несколько самцов. Самцы спариваются с несколькими самками (полигиния), самки спариваются с несколькими самцами (полиандрия). Вследствие этого в одном помете могут появиться полуродные братья и сестры, однако, тех крысят, чьим отцом будет являеться доминантный самец, все же родится больше. Такое беспорядочное спаривание особей с разными партнерами называется промискуитет.
Итак, при низкой плотности населения самцы серых крыс территориальны и полигамны; при высокой плотности отношения между особями внутри группы регулируются системой социальной иерархии, система спаривания типа промискуитет.
Агонистическое поведение
Иерархические отношения между половозрелыми самцами в колонии устанавливаются в результате их борьбы за доминирующее положение, которая может иметь вид драки с нанесением физических повреждений или оканчиваться очень быстро. Однако, после того как эти отношения уже установлены, частота и выраженность конфликтов становится незначительной. Доминантом, как правило, становится более крупный самец. Что же касается иерархических отношений между подчиненными особями, то они до сих пор не ясны. Известно, что крысы обоих полов с низким социальным статусом живут в среднем меньше, чем крысы с более высоким рангом. Причина – социальный стресс*, который испытывают первые. Не будем забывать, что и без этого до своего первого дня рождения доживает лишь 5% диких крыс.
Чаще агонистическое поведение можно наблюдать у самцов, достигших шестимесячного возраста. Надо также заметить, что агрессивные столкновения у декоративных крыс, живущих в клетках, длятся дольше, чем у диких, а также у вторых
отсутствуют некоторые виды агонистического поведения (например, переворачивание на спину), которые можно наблюдать у домашних крыс.
Иерархия
В колонии подчиняющиеся самцы используют две стратегии в их отношении с доминирующим самцом: одни (омега-самцы) избегают самца-доминанта, другие (бета-самцы) стремятся постоянно быть рядом с ним.
Когда омега-самцы встречаются с доминирующим самцом, они вызывают у него агрессию. Такие самцы в естественных условиях, как правило, уходят из колонии. Однако, если убрать из колонии главного самца, новым доминантом зачастую становится именно омега-самец. Бета-самцы в такой ситуации становятся полностью покорными новой доминирующей крысе, та, в свою очередь, не возражает против их присутствия.
Таким образом, обе стратегии имеют свои плюсы и минусы. Беты живут в относительном мире с доминирующим самцом и имеют больший доступ к пище и самкам. Кто-то из омег, с другой стороны, может при случае стать новым альфа-самцом (доминантом).
Выбор крысой одной из стратегий зависит от ее психофизиологических характеристик*.
Пояснение: доминирующим может называться такое животное, которое способно действовать (при половом, агрессивном, пищевом и другом поведении), не считаясь с действиями своих партнеров. Соответственно, подчиненное животное – то, чьи действия вызываются, модифицируются, ограничиваются или подавляются действиями партнеров.
Аллогруминг
Аллогруминг – чистка меха одной особи другой особью. Взаимные чистки широко распространены у млекопитающих, ведущих семейно-групповой образ жизни, в том числе и у крыс. Аллогруминг играет важную роль в поддержании системы социальных связей в группе, в подавлении агрессивных тенденций. Аллогруминг оказывает умиротворяющее воздействие на партнера, вызывая ощущение удовольствия. Одна особь чистит мех у другой особи в тех местах, до которых та не может достать своей мордочкой при автогруминге; аллогруминг также играет роль в создании общего
«запаха колонии». Самки большинства видов млекопитающих вылизывают и чистят мех у детенышей (у моногамных видов или факультативно моногамных видов (джунгарский хомячок) это проделывает и самец). У крыс и других грызунов с семейно-групповым образом жизни (например, монгольская и большая песчанки) существуют особые позы «подставления под чистку», когда соподчиненная особь ищет тактильного контакта с доминантом.
Метки
Учеными обнаружено, что в моче крыс содержатся не только продукты обмена веществ, но целый ряд компонентов – феромонов, которые у крыс служат сигналами, определяющими положение и состояние индивидуума. Поэтому животные оставляют в пределах своего участка обитания капельки мочи (метки), чтобы обозначить пределы своей территории, а также с целью привлечения половых партнеров.
Легко заметить, когда крыса метится: она прижимает заднюю часть тела к «земле» и идет заметно медленнее, чем обычно. Под светом ультрафиолетовой лампы (см. ratmania.narod.ru/ultra.html)
отчетливо видны даже старые метки (стоит отметить, что крысы могут видеть часть лучей ультрафиолетового спектра света).
Подчиненные особи и самки метятся относительно редко и не сильно. Однако, в ночь перед течкой самка, чтобы заявить о своей готовности к спариванию, начинает помечать предметы чаще, чем обычно.
Такие мочевые метки - не единственный вид обонятельной коммуникации у крыс. Иногда крысы трутся о вертикальные поверхности (например, о стену) или о край входа в нору, чтобы оставить на них секреты специфических желез, расположенных на боках их тела. Самцы делают это чаще, чем самки (также крысы чаще трутся о поверхности, если чувствуют запах других крыс).
Пища
Вопрос, как крысы узнают, что можно есть, а что нельзя, хорошо изучен. Крысы всеядны, т.е. они могут есть почти все. Такая способность есть что угодно делает крыс очень гибкими в плане приспосабливаемости. Вследствие этого их среды обитания могут быть очень разнообразными. Поскольку
крысы едят не какой-либо определенный вид корма, они должны уметь различать полезную пищу от вредной и опасной. Как же молодые крысы различают хорошую пищу от опасной?
Удивительно, но даже эмбрионы узнают, что съела вынашивающая их самка, и в будущем предпочитают есть те корма, которые она употребляла во время беременности. Далее, когда крысята уже родились, через молоко матери они получают информации о том, чем она питалась. Перейдя на самостоятельное питание, они будут уверенно есть ту же пищу.
Подросшие крысята, встречая незнакомый вид корма, следят, едят ли его взрослые крысы колонии. Если пища помечена взрослой крысой, значит, ее можно есть не опасаясь.
Немалую роль в выборе пищи играет и жизненный опыт. Крыса сначала пробует небольшой кусочек от неизвестной пищи, и если после этого она почувствует себя плохо, то больше она ее есть не будет.
Следует также отметить, что крысы обладают хорошо развитыми чувствами обоняния и вкуса, поэтому нередко они определяют,
опасна ли пища, даже не попробовав или не проглотив ее.
А что если крыса все-таки съела ядовитую пищу?
Крысы не способны отрыгивать съеденную пищу. Вместо этого, почувствовав тошноту, они ищут и поедают такие вещества, как глину, грязь, а иногда и подстилку гнезда. Эти вещества нейтрализуют токсины, которые и вызывают тошноту.
Неспособность крыс отрыгивать пищу не является примитивной чертой их организации. Все предки грызунов, вплоть до рыб, обладали такой способностью.
Значение капрофагии за время перехода крысенка на твердые корма
В формировании способности крысенка к перевариванию твердых кормов большое значение имеет поедание экскрементов матери, привлекательность которых для него резко повышается с двухнедельного возраста.
Оказывается, через 14 дней после родов у самки начинается секреция специального феромона, вызывающего у детенышей соответствующую реакцию. Максимальную реакцию проявляют детеныши 21-дневного возраста, у 27-дневных крысят реакция
резко ослабевает (у самки прекращается секреция феромона).
Как объем, так и состав фекальных масс самки, когда крысята достигают двухнедельного возраста, сильно изменяются. Вес фекальных масс в этот период в 2 – 3 раза превышает таковой девственных самок. Соответственно увеличивается и потребление ими кормов. В фекалиях резко повышается содержание летучих компонентов, они светлее обычных, сильнее пахнут и менее структурированы.
Оказывается, увеличение уровня пролактина в крови самки приводит к постепенному увеличению концентрации этого гормона в печени, а это, в свою очередь, приводит к увеличению синтеза печенью основной желчной – холиевой кислоты. В слепой кишке крыс под воздействием определенных бактерий из холиевой кислоты синтезируется вторичная желчная – деоксихолиевая кислота. Потребность в ней и заставляет крысят поедать экскременты матери, а привлекает их феромон-содержащий компонент фекалий – производное диоксихолиевой кислоты.
Вплоть до 28-дневного возраста крысят уровень секреции деоксихолиевой кислоты у них значительно ниже, чем у взрослых крыс.
Потребность в деоксихолиевой кислоте может объясняться по крайней мере двумя основными причинами. Первая - повышение иммунозащиты у крысят, вторая – миелинизация мозга.
Кроме того, можно предположить, что фекалии матери обогащены и кишечной микрофлорой, специфичной для предпочитаемых ею кормов, т.е. капрофагия также способствует физиологической пищевой специализации крысят.
Норы
В природе дикие крысы живут в норах. Крысиная нора может представлять собой гнездовую камеру, соединенную с поверхностью земли коротким туннелем (в самом простом случае), или довольно сложную систему туннелей, проходов и камер.
Размер и структура норы зависит от размера и структуры колонии, использующей эту нору в качестве убежища. Если число особей в колонии вследствие каких-либо причин резко падает, то оставшиеся крысы уже не в состоянии ухаживать за всей норой,
ее состояние сильно ухудшается.
Вход в нору обычно располагается в защищенном или невидимом для хищников месте. Крысы могут закрывать вход пробкой из травы или земли. Средний диаметр туннеля составляет 8.3 см, средняя длина прямого туннеля – около 30 см (в конце туннель поворачивает в другую сторону, раздваивается или оканчивается тупиком или камерой). Камеры сильно варьируют в размерах. Средняя камера размером 18 на 22 см может вместить 7 крыс. Бывают очень маленькие камеры, способные вместить только трех крыс, или очень большие – в них могут находиться сразу 11 крыс.
Вне норы крысы предпочитают перемещаться по определенным системам троп – это позволяет им быстро сориентироваться в случае опасности и спрятаться в норе или другом убежище.
- По материалам сайта "Rat behavior and biology"
- П о материалам книги "Серая крыса. Систематика, экология, регуляция численности" (Ред. В.Е.Соколов, Е.В.Карасева)
Клеточные культуры
Мышь
Нематода
Почвенная нематода - Удобный объект исследования развития и поведения.
короткий жизненный цикл - 3 дня, малый размер (1,5 мм в длинну),
тысячи нематод могут одновременно жить в чашках петри на газонах с кишечной палочкой Escherichia coli.
Одна нематода производит 300-350 потомков.
C.elegans - гермафродиты, способные к самовоспроизводству или могут спариваться с самцами.
постоянное и небольшое количество клеток 1000 штук, включая 302 клетки нервной системы у гермафродитов и 381 у самцов. Были установлены все связи между нервными клетками и определена роль каждого гена вовлеченного в развитие и функционирование нервной системы.
Геном всего в 20 раз больше чем у кишечной палочки.
Развитие червя включает 14 часов эмбриогенеза и 36 часов постэмбрионального развития, включающего L1-L4 стадии и взрослое состояние.
Гермафродиты начинают вырабатывать сперму на стадии L4 и яйцеклетки во взрослом состоянии. Взрослые нематоды - самки, хранящие ранее произведенную сперму в спермтеке. Гермафродиты производят яйцеклеток на много больше чем сперматозоидов.
X0 самцы появляются спонтанно из ХХ гермафродитной популяции при нерасхождении Х хромосом с частотой 0,1%. Гермафродиты спаривающиеся с самцамии производят самцов и и гермафродитов в соотношении 1:1. Хотя это соотношение сдвинуто больше в сторону гермафродитов.
Анатомически все нематоды представляют собой две концентрические трубки разделенные пространством - целомом, наполненном жидкостью. Наружная трубка покрыта коллагеновой внеклеточной кутикулой, вырабатываемой подлежащей гиподермисом.
Во время каждой из четырех линек старая кутикула сбрасывается и образуется новая. Мускулатура тела представлена четырмя продольными тяжами прикрепленными к кутикуле через гиподермис. Сокращение субвентральных мышечных тяжей и расслабление субдорсальных и наоборот приводит к синусоидальным движениям в дорсо-вентральной плоскости.
Нервная система, гонады, выделительная система располагаются внутри целома.
Внутренняя трубка состоит из мышечного фаринкса с автономной нервной системой и кишечника.
Бластомер обрзуется при асимметричном асинхронном делении яйцеклетки, в результате чего образуется четыре предшественника соматических клеток и одна клетка зародышевого пути. Эмбрион производит 671 клетку, 113 из которых подвергаются апоптозу. 10% из 558 образованных клеток делятся дальше образуя взрослую нематоду.
Фаги
Фаг - λ
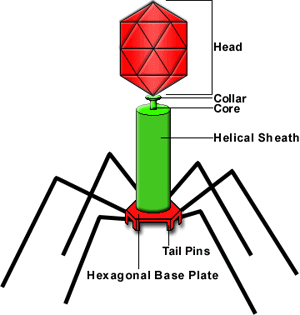 рис.1 Внешнее строение фага - λ.
рис.1 Внешнее строение фага - λ.
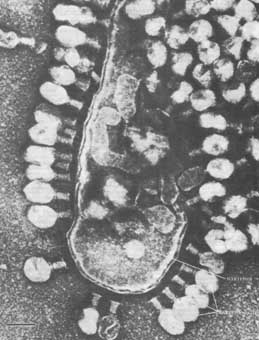 рис. Фаг - λ поражающий E.coli.
рис. Фаг - λ поражающий E.coli.