Гетерохроматин
ДНК способна связываться с различными белками, образуя структуру называемую хроматином. Хроматин подразделяется на два функциональных состояния: эухроматин и гетерохроматин. Гетерохроматин разделяется на конститутивный - существующий постоянно в течении всего клеточного цикла и факультативный, образующийся в определенные моменты времени в определенных местах. К отличительным свойствам гетерохроматина относят: конденсированное состояние в интерфазе, транскрипционно неактивное состояние, поздняя репликация в S-фазе клеточного цикла ... Так же к особенностям гетерохроматина можно отнести недорепликацию в политенных хромосомах у дрозофилы.
Эухроматин обогащен следующими модификациями: ацетилирование H3 и H4, метилирование H4K20, H4K9 и H3K27.
Для гетерохроматина характерно метилирование H3K9, но эта модификация встречается также и в эухроматине, особенно в ORF.
Цитологическое строение гетерохроматина
Формирование гетерохроматина у Дрозофилы
Наиболее ранний этап формирования гетерохроматина (ГХ) происходит при участии механизма интерференции РНК, при котором sRNA (small RNAs) воздействуют на места будущего гетерохроматина посредством RITS комплекса (см. обзор РНК интерференция).
На следующей стадии происходит замена гистона H2A гистоном H2Av, что запускает процессы ацетилирования H4K12 и деацетилирование H3K9 (см. обзор Гистоны). Деацетилированный Н3K9 метилируется ферментом Su(var)3-9.
К модифицированному таким образом гистону присоединяется HP1 (heterochromatin protein 1), способствующий присоединению гистон метилтрансферазы Suv4-20, триметилирующей гистон H4K20.
Таким образом образование гетерохроматина проходит по следующей схеме
замена H2A на H2Av--> действие RITS-комплекса--> ацетилирование H4K12 --> деацетилирование и метелирование H3K9 --> связывание Me-H3K9 белка HP1 --> метилирование H4K20.
Формирование гетерохроматина у дрожжей
У Sp наблюдается классический эффект положения гена (см. обзор Эффект положения гена) в пределах центромер, теломер и локуса ответственного за определение пола, что говорит о наличии гетерохроматина. Необходимо отметить, что хромосомы дрожжей, в отличие от политенных хромосом дрозофилы, не видны в световой микроскоп, поэтому выявить гетерохроматиновые участки можно только косвенным путем.
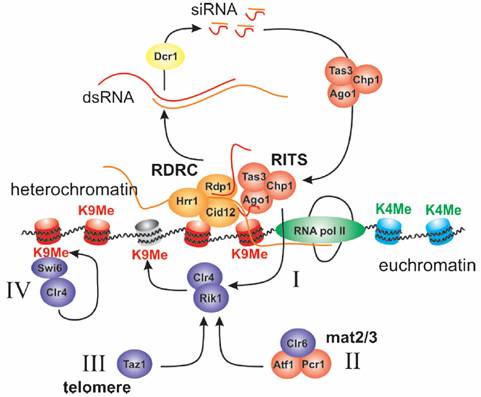 рис.1 Механизмы, способствующие образованию гетерохроматина в различных местах.
рис.1 Механизмы, способствующие образованию гетерохроматина в различных местах.
Образованию гетерохроматина предшествует метилирование H3K9 с последующим присоединением HP1 (Swi6). Конвергентные промоторы в пределах повторов центромерной ДНК образуют dsRNA, которые Dcr1 превращает в siRNA. siRNA связываются с RITS и атакуют образующиеся в промоторах транскрипты, связываясь с ними через комплиментарные взаимодействия. RITS и RDRC комплексы объединяются, увеличивая действие сигнала и стабилизируя процесс. Такой RNAi зависимый процесс основной в установлении метилирования H3K9 (механизм I). Локус ответственный за определение пола определяется HDAC связывания через белки связывающие ДНК (механизм II), усиливая этот процессс, тогда как в теломерах Taz1-зависимый мханизм уменьшается RNAi (механизм III). После метилирования H3K9 белки Clr4 и Swi6 связываются с хромосомой, поддерживая MeH3K9 эпигенетически (механизм IV).
Гены в гетерохроматине
 Гены в гетерохроматине L-плеча второй хромосомы дрозофилы
Гены в гетерохроматине L-плеча второй хромосомы дрозофилы
Сокращения.
H4K12 - так обозначается определенная аминокислота в гистоновых белках. В данном случае это лизин (K) в девятом положении гистона H4.
ORF (Open Reading Frames) - рамка считывания, участок ДНК с которого считывается РНК.
Sp (Schizosaccharomyces pombe) - вид дрожжей
Механизмы диминуции
Привет, Настя!
Литература:
- Swaminathan J., Baxter E., Corces V. (2005) The role of histone H2Av variant replacement and histone H4 acetylation in the establishment of Drosophila heterochromatin. Genes & development 19:65-76.
- Verdel A, Jia S., Gerber S., Sugiyama T., Gygi S., Grewal S., Moazed D. (2004) RNAi-mediated targeting of heterochromatin by the RITS complex. Science, 30;303(5658):672-6.
- HornP. J., Peterson C.L. (2006) Heterochromatin assembly: A new twist on an old model. Chromosome Research 14:83Y94
- Fransz P. ,Hoopen R.,Tessadori F. (2006) Composition and formation of heterochromatin in Arabidopsis thaliana. Chromosome Research 14:71–82