Аппарат Гольджи
 Аппарат Гольджи
Аппарат Гольджи
Эндоплазматический ретикулум, плазматическая мембрана и аппарат Гольджи составляют единую мембранную систему клетки, в пределах которой происходят процессы обмена белками и липидами с помощью направленного и регулируемого внутриклеточного мембранного транспорта.
Каждая из мембранных органелл характеризуется уникальным составом белков и липидов.
Строение АГ
АГ состоит из группы плоских мембранный мешков - цистерны, собранные в стопки - диктиосомы (~5-10 цистерн, у низших эукариот >30). Число диктиосом в разных клетках от 1 до ~500.
Отдельные цистерны диктиосомы переменной толщины - в центре ее мембраны сближены - просвет 25 нм, на переферии образуются расширения - ампулы ширина которых не постоянна. От ампул отшнуровываются ~50нм-1мкм пузырьки связанные с цистернами сетью трубочек.
У многоклеточных организмов АГ состоит из стопок цистерн связанных между собой в единую мембранную систему. АГ представляет собой полусферу, основание которой обращено к ядру. АГ дрожжей представлен изолированными единичными цистернами, окруженными мелкими пузырьками, тубулярной сетью, секреторными везикулами и гранулами. У мутантов дрожжей Sec7 и Sec14 наблюдается структура, напоминающая стопку цистерн клеток млекопитающих.
Для АГ характерна полярность его структур. Каждая стопка имеет два полюса: проксимальный полюс (формирующийся, цис-поверхность) и дистальный (зрелый,
транс-поверхность). Цис-полюс – сторона мембраны с которой сливаются пузырьки. Транс-полюс – сторона мембраны от которой пузырьки отпочковываются.
Пять функциональных компартментов АГ:
1. Промежуточные везикуло-тубулярные структуры (VTC или ERGIC - ER-Golgi intermediate compartment)
2. Цис-цистерна (cis) - цистерны расп ближе к ЭР:
3. Срединные (medial) цистерны – центральные цистерны
4. Транс-цистерна (trans) - наиболее удаленные от ЭР цистерны.
5. Тубулярная сеть, примыкающая к трансцистерне - транссеть Гольджи (TGN)
Стопки цистерн изогнуты, так что вогнутая трансповерхность обращена к ядру.
В среднем в АГ 3-8 цистерн, в активно секретирующих клеток может быть больше (в экзокринных клетках поджелудочной железы до 13).
Каждая цистерна имеет цис и транс поверхности. Синтезированные белки, мембранные липиды, гликозилированные в ЭР, попадают в АГ через цис-полюс. Вещества через стопки передаются транспортными
пузырьками отделяющиеся от ампул. При прохождении белков или липидов через стопки Гольджи, они претерпевают серию посттрансляционных модификаций, включающих изменение N-связанных олигосахаридов:
цис: маннозидазаI подравнивает длинные маннозные цепи до М-5
промежуточный: N-ацетилглюкоэаминтрансферазаI переносит N-ацетилглюкозамин
транс: добавляются концевые сахара –остатки галактозы и сиаловая к-та.
Строение Аппарата Гольджи и схема транспорта.
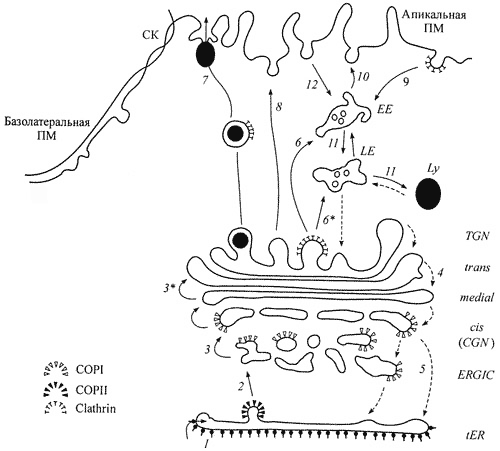 Пять компанентов АГ и схема транспорта: промежуточный (ERGIC), цис, промежуточный, транс и транссеть Гольджи (TGN). 1. Вход синтезированных белков, мембранных гликопротеинов и лизосомных ферментов в цистерну переходного ЭР, прилегающую к АГ и 2 - их выход из ЭР в пузырьках окаймленных COPI (антероградный транспорт). 3 - возможный транспорт карго от тубуло-везикулярных
Пять компанентов АГ и схема транспорта: промежуточный (ERGIC), цис, промежуточный, транс и транссеть Гольджи (TGN). 1. Вход синтезированных белков, мембранных гликопротеинов и лизосомных ферментов в цистерну переходного ЭР, прилегающую к АГ и 2 - их выход из ЭР в пузырьках окаймленных COPI (антероградный транспорт). 3 - возможный транспорт карго от тубуло-везикулярных
кластеров к цис-цистерне АГ в пузырьках COPI; 3* - транспорт карго от более ранних к более поздним цистернам; 4 - возможный ретроградный везикулярный транспорт карго между цистернами АГ; 5 - возврат резидентных протеинов из АГ в tER с помощью пузырьков, окаймленных COPI (ретроградный транспорт); 6 и 6* - перенос лизосомных ферментов с помощью окаймленных клатрином пузырьков соответственно в ранние EE и поздние LE эндосомы; 7 - регулируемая секреция секреторных гранул; 8 - конститутивное встраивание мембранных белков в апикальную плазматическую мембрану ПМ; 9 - опосредованный рецептором эндоцитоз с помощью окаймленных клатрином пузырьков; 10 возвращение ряда рецепторов из ранних эндосом в плазматическую мембрану; 11 - транспорт лигандов из EE в LE и и лизосомы Ly; 12 - транспорт лигандов в неклатриновых пузырьках.
Функции АГ
1. Транспорт - через АГ проходят три группы белков: белки периплазматической мембраны, белки, предназначенные
на экспорт из клетки, и лизосомные ферменты.
2. Cортировка для транспорта: сортировка для дольнейшего транспорта к органеллам, ПМ, эндосомам, секреторным пузырькам происходит в транс-комплексе Гольджи.
3. Секреция - секреция продуктов, синтезируемых в клетке.
3. Гликозилирование белков и липидов: гликозидазы удаляют остатки сахаров - дегликозилирование, гликозилтрансферазы прикрепляют сахара обратно на главную углеводную цепь - гликозилирование.В нем происходят гликозилирование олигосахаридных цепей белков и липидов, сульфатирование ряда ахаров и тирозиновых остатков белков, а также активация предшественников полипептидных гормонов и нейропептидов.
4. Синтез полисахаридов - многие полисахариды образуются в АГ в том числе пектин и гемицеллюлоза, образующие клеточные стенки растений и большинство гликозаминогликанов образующих межклеточный матрикс у животных
5. Сульфатирование - большинство сахаров, добавляемых к белковай сердцевине протеогликана, сульфатируются
6. Добавление маннозо-6-фосфата: М-6-P добавляется как направляюций сигнал к ферментам, предназначенным для лизосом.
ГЛИКОЗИЛИРОВАНИЕ
Большинство белков начинает гликозилироваться в шероховатом ЭР посредством добавления к растущей полипептидной цепи N-связанных олигосахаридов. Если гликопротеин свернут в нужной конформации, он выходит из ЭР и направляется в АГ, где происходит его посттрансляционная модификация.
В гликозилировании секретируемых продуктов принимают участие ферменты - гликозилтрансферазы. Они участвуют в ремоделированиии Т-связанных боковых олигосахаридных цепей и добвлении О-связанных гликанов и олигосахаридных частей протеогликанов гликолипидов.В модификации олигосахаридов участвуют фрменты а-маннозидаза I и II, которые также являются резидентными белками АГ.
Кроме того в АГ происходит гликозилирование липидно-протеиновых мембранных доменнов, называемых рафтами.
Долихолфосфат добавляет углеводный комплекс – 2GlcNAc-9-манноз-3-глюкозы к аспарагину растущего полипептида. Терминальная глюкоза отщепляется в два этапа: глюкозидаза I отщепляет терминальный остаток глюкозы, глюкозидаза II удаляет еще два остатка глюкозы. Затем отщепляется манноза. На этом начальный этап процессинга углеводов в ЭР завершается и белки несущие олигосахаридный комплекс, поступают в АГ
В первых цистернах АГ удаляются еще три остатка маннозы. На этой стадии стержневой комплекс имеет еще 5 маннозных остатков. N-ацетилглюкозаминтрансфераза I добавляет один остаток N-ацетилглюкозамина GlcNAc. От образовавшегося комплекса отщепляется еще 3 остатка маннозы. Состоит теперь из двух молоекул GlcNAc-3-маннозо-1-GlcNAc является стержневой структурой, к которой гликозилтрансферезы добавляют другие
углеводы. Каждая гликозилтрансфераза распознает развивающуюся углеводную структуру и добавляет к цепи свой собственный сахарид.
СЕКРЕЦИЯ
Схема секреции:
Синтезированные в ЭР белки концентрируются в сайтах выхода переходного ЭР благодаря активности коатомерного комплекса COPII и сопутствующих компонентов и транспортируются в промежуточный между ЭР и АГ компартмент ERGIC, из которого они переходят в АГ в отпочковывающихся пузырьках, или по тубулярным структурам. Белки ковалентно модифицируются, проходя через цистерны АГ, на транс-поверхности АГ сортируются и отправляются к местам своего назначения. Секреция белков требует пассивного встраивания новых мембранных компонентов в плазматическую мембрану. Для восстановления баланса мембран служит контитутивный рецепторопосредованный эндоцитоз.
Эндо и экзоцитозный пути переноса мембран имеют общие закономерности в направленности движения мембранных переносчиков к сооответствующей
мишени и в специфичности слияния и почкования. Основным местом встречи этих путей является АГ.