Пластиды
 Хлоропласт
Хлоропласт
Пластиды – это мембранные органоиды, встречающиеся у фотосинтезирующих эукариотических организмов (высшие растения, низшие водоросли, некоторые одноклеточные организмы). Пластиды окружены двумя мембранами, в их матриксе имеется собственная геномная система, функции пластид связаны с энергообеспечением клетки, идущим на нужды фотосинтеза.
У высших растений найден целый набор различных пластид (хлоропласт, лейкопласт, амилопласт, хромопласт), представляющих собой ряд взаимных превращений одного вида пластиды в другой. Основной структурой, которая осуществляет фотосинтетические процессы, является хлоропласт.
У высших растений также встречается деление зрелых хлоропластов, но очень редко. Увеличение числа хлоропластов и образование других форм пластид (лейкопластов и хромопластов) следует рассматривать как путь превращения структур-предшественников, пропластид. Весь же процесс развития различных пластид можно представить в виде монотропного (идущего в одном направлении) ряда смены форм:
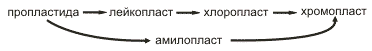
Многими исследованиями был установлен необратимый характер онтогенетических переходов пластид. У высших растений возникновение и развитие хлоропластов происходят через изменения пропластид. Пропластиды представляют собой мелкие (0,4-1 мкм) двумембранные пузырьки, не имеющие отличительных черт их внутреннего строения. Они отличаются от вакуолей цитоплазмы более плотным содержимым и наличием двух отграничивающих мембран, внешней и внутренней. Внутренняя мембрана может давать небольшие складки или образовывать мелкие вакуоли. Пропластиды чаще всего встречаются в делящихся тканях растений (клетки меристемы корня, листьев, в точки роста стеблей и др.). По всей вероятности, увеличение их числа происходит путем деления или почкования, отделения от тела пропластиды мелких двумембранных пузырьков.
Хлоропласты
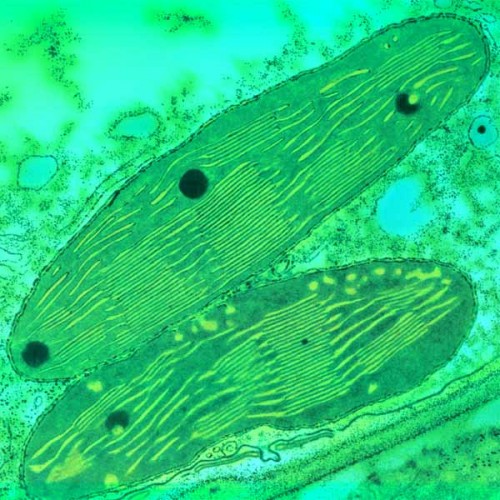 Хлоропласты – это структуры, в которых происходят фотосинтетические процессы, приводящие в конечном итоге к связыванию углекислоты, к выделению кислорода и синтезу сахаров. структуры удлиненной формы с шириной 2-4 мкм и протяженностью 5-10 мкм. У зеленых водорослей встречаются гигантские хлоропласты (хроматофоры), достигающие длины 50 мкм.
Хлоропласты – это структуры, в которых происходят фотосинтетические процессы, приводящие в конечном итоге к связыванию углекислоты, к выделению кислорода и синтезу сахаров. структуры удлиненной формы с шириной 2-4 мкм и протяженностью 5-10 мкм. У зеленых водорослей встречаются гигантские хлоропласты (хроматофоры), достигающие длины 50 мкм.
у зеленых водорослей может быть по одному хлоропласту на клетку. Обычно на клетку высших растений приходится в среднем 10-30 хлоропластов. Встречаются клетки с огромным количеством хлоропластов. Например, в гигантских клетках палисадной ткани махорки обнаружено около 1000 хлоропластов.
Хлоропласты представляют собой структуры, ограниченные двумя мембранами – внутренней и внешней. Внешняя мембрана, как и внутренняя, имеет толщину около 7 мкм, они отделены друг от друга межмембранным пространством около 20-30 нм. Внутренняя мембрана хлоропластов отделяет строму пластиды, аналогичную матриксу митохондрий. В строме зрелого хлоропласта высших растений видны два типа внутренних мембран. Это – мембраны, образующие плоские, протяженные ламеллы стромы, и мембраны тилакоидов, плоских дисковидных вакуолей или мешков.
Ламеллы стромы (толщиной около 20 мкм) представляют собой плоские полые мешки или же имеют вид сети из разветвленных и связанных друг с другом каналов, располагающихся в одной плоскости. Обычно ламеллы стромы внутри хлоропласта лежат параллельно друг другу и не образуют связей между собой.
Кроме мембран стромы в хлоропластах обнаруживаются мембранные тилакоиды. Это плоские замкнутые мембранные мешки, имеющие форму диска. Величина межмембранного пространства у них также около 20-30 нм. Такие тилакоиды образуют стопки наподобие столбика монет, называемые гранами.
Число тилакоидов на одну грану очень варьирует: от нескольких штук до 50 и более. Размер таких стопок может достигать 0,5 мкм, поэтому граны видны в некоторых объектах в световом микроскопе. Количество гран в хлоропластах высших растений может достигать 40-60. Тилакоиды в гране сближены друг с другом так, что внешние слои их мембран тесно соединяются; в месте соединения мембран тилакоидов образуется плотный слой толщиной около 2 нм. В состав граны кроме замкнутых камер тилакоидов обычно входят и участки ламелл, которые в местах контакта их мембран с мембранами тилакоидов тоже образуют плотные 2-нм слои. Ламеллы стромы, таким образом, как бы связывают между собой отдельные граны хлоропласта. Однако полости камер тилакоидов всезда замкнуты и не переходят в камеры межмембранного пространства ламелл стромы. Ламеллы стромы и мембраны тилакоидов образуются путем отделения от внутренней мембраны при начальных этапах развития пластид.
В матриксе (строме) хлоропластов обнаруживаются молекулы ДНК, рибосомы; там же происходит первичное отложение запасного полисахарида, крахмала, в виде крахмальных зерен.
Характерным для хлоропластов является наличие в них пигментов, хлорофиллов, которые и придают окраску зеленым растениям. При помощи хлорофилла зеленые растения поглощают энергию солнечного света и превращают ее в химическую.
Функции хлоропластов
Геном пластид
Подобно митохондриям, хлоропласты имеют собственную генетическую систему, обеспечивающую синтез ряда белков внутри самих пластид. В матриксе хлоропластов обнаруживаются ДНК, разные РНК и рибосомы. Оказалось, что ДНК хлоропластов резко отличается от ДНК ядра. Она представлена циклическими молекулами длиной до 40-60 мкм, имеющими молекулярный вес 0,8-1,3х108 дальтон. В одном хлоропласте может быть множество копий ДНК. Так, в индивидуальном хлоропласте кукурузы присутствует 20-40 копий молекул ДНК. Длительность цикла и скорость репликации ядерной и хлоропластной ДНК, как было показано на клетках зеленых водорослей, не совпадают. ДНК хлоропластов не состоит в комплексе с гистонами. Все эти характеристики ДНК хлоропластов близки к характеристикам ДНК прокариотических клеток. Более того, сходство ДНК хлоропластов и бактерий подкрепляется еще и тем, что основные регуляторные последовательности транскрипции (промоторы, терминаторы) у них одинаковы. На ДНК хлоропластов синтезируются все виды РНК (информационная, трансферная, рибосомная). ДНК хлоропластов кодирует рРНК, входящую в состав рибосом этих пластид, которые относятся к прокариотическому 70S типу (содержат 16S и 23S рРНК). Рибосомы хлоропластов чувствительны к антибиотику хлорамфениколу, подавляющему синтез белка у прокариотических клеток.
Так же как в случае хлоропластов мы вновь сталкиваемся с существованием особой системы синтеза белка, отличной от таковой в клетке.
Эти открытия вновь пробудили интерес к теории симбиотического происхождения хлоропластов. Идея о том, что хлоропласты возникли за счет объединения клеток-гетеротрофов с прокариотическими синезелеными водорослями, высказанная на рубеже XIX и XX вв. (А.С. Фоминцин, К.С.Мережковский) вновь находит свое подтверждение. В пользу этой теории говорит удивительное сходство в строении хлоропластов и синезеленых водорослей, сходство с основными их функциональными особенностями, и в первую очередь со способностью к фотосинтетическим процессам.
Известны многочисленные факты истинного эндосимбиоза синезеленых водорослей с клетками низших растений и простейших, где они функционируют и снабжают клетку-хозяина продуктами фотосинтеза. Оказалось, что выделенные хлоропласты могут также отбираться некоторыми клетками и использоваться ими как эндосимбионты. У многих беспозвоночных (коловратки, моллюски), питающихся высшими водорослями, которые они переваривают, интактные хлоропласты оказываются внутри клеток пищеварительных желез. Так, у некоторых растительноядных моллюсков в клетках найдены интактные хлоропласты с функционирующими фотосинтетическими системами, за активностью которых следили по включению С14О2.
Как оказалось, хлоропласты могут быть введены в цитоплазму клеток культуры фибробластов мыши путем пиноцитоза. Однако они не подвергались атаке гидролаз. Такие клетки, включившие зеленые хлоропласты, могли делиться в течение пяти генераций, а хлоропласты при этом оставались интактными и проводили фотосинтетические реакции. Были предприняты попытки культивировать хлоропласты в искусственных средах: хлоропласты могли фотосинтезировать, в них шел синтез РНК, они оставались интактными 100 ч, у них даже в течение 24 ч наблюдались деления. Но затем происходило падение активности хлоропластов, и они погибали.
Эти наблюдения и целый ряд биохимических работ показали, что те черты автономии, которыми обладают хлоропласты, еще недостаточны для длительного поддержания их функций и тем более для их воспроизведения.
В последнее время удалось полностью расшифровать всю последовательность нуклеотидов в составе циклической молекулы ДНК хлоропластов высших растений. Эта ДНК может кодировать до 120 генов, среди них: гены 4 рибосомных РНК, 20 рибосомных белков хлоропластов, гены некоторых субъединиц РНК-полимеразы хлоропластов, несколько белков I и II фотосистем, 9 из 12 субъединиц АТФ-синтетазы, части белков комплексов цепи переноса электронов, одной из субъединиц рибулозодифосфат-карбоксилазы (ключевой фермент связывания СО2), 30 молекул тРНК и еще 40 пока неизвестных белков. Интересно, что сходный набор генов в ДНК хлоропластов обнаружен у таких далеко отстоящих представителей высших растений как табак и печеночный мох.
Основная же масса белков хлоропластов контролируется ядерным геномом. Оказалось, что ряд важнейших белков, ферментов, а соответственно и метаболические процессы хлоропластов находятся под генетическим контролем ядра. Так, клеточное ядро контролирует отдельные этапы синтеза хлорофилла, каротиноидов, липидов, крахмала. Под ядерным контролем находятся многие энзимы темновой стадии фотосинтеза и другие ферменты, в том числе некоторые компоненты цепи транспорта электронов. Ядерные гены кодируют ДНК-полимеразу и аминоацил-тРНК-синтетазу хлоропластов. Под контролем ядерных генов находится большая часть рибосомных белков. Все эти данные заставляют говорить о хлоропластах, так же как и о митохондриях, как о структурах с ограниченной автономией.
Транспорт белков из цитоплазмы в пластиды происходит в принципе сходно с таковым у митохондрий. Здесь также в местах сближения внешней и внутренней мембран хлоропласта располагаются каналообразующие интегральные белки, которые узнают сигнальные последовательности хлоропластных белков, синтезированных в цитоплазме, и транспортируют их в матрикс-строму. Из стромы импортируемые белки согласно дополнительным сигнальным последовательностям могут включаться в мембраны пластиды (тилакоиды, ламеллы стромы, внешняя и внутренняя мембраны) или локализоваться в строме, входя в состав рибосом, ферментных комплексов цикла Кальвина и др.
Удивительное сходство структуры и энергетических процессов у бактерий и митохондрий, с одной стороны, и у синезеленых водорослей и хлоропластов – с другой, служит веским аргументом в пользу теории симбиотического происхождения этих органелл. Согласно этой теории, возникновение эукариотической клетки прошло через несколько этапов симбиоза с другими клетками. На первой стадии клетки типа анаэробных гетеротрофных бактерий включили в себя аэробные бактерии, превратившиеся в митохондрии. Параллельно этому в клетке-хозяине прокариотический генофор формируется в обособленное от цитоплазмы ядро. Так могли возникнуть гетеротрофные эукариотические клетки. Повторные эндосимбиотические взаимоотношения между первичными эукариотическими клетками и синезелеными водорослями привели к появлению в них структур типа хлоропластов, позволяющих клеткам осуществлять автосинтетические процессы и не зависеть от наличия органических субстратов (рис. 236). В процессе становления такой составной живой системы часть генетической информации митохондрий и пластид могла изменяться, перенестись в ядро. Так, например две трети из 60 рибосомных белков хлоропластов кодируется в ядре и синтезируются в цитоплазме, а потом встраивается в рибосомы хлоропластов, имеющие все свойства прокариотических рибосом. Такое перемещение большой части прокариотических генов в ядро привело к тому, что эти клеточные органеллы, сохранив часть былой автономии, попали под контроль клеточного ядра, определяющего в большей степени все главные клеточные функции.
Пропластиды
При нормальном освещении пропластиды превращаются в хлоропласты. Сначала они растут, при этом происходит образование продольно расположенных мембранных складок от внутренней мембраны. Одни из них простираются по всей длине пластиды и формируют ламеллы стромы; другие образуют ламеллы тилакоидов, которые выстраиваются в виде стопки и образуют граны зрелых хлоропластов. Несколько иначе развитие пластид происходит в темноте. У этиолированных проростков происходит в начале увеличение объема пластид, этиопластов, но система внутренних мембран не строит ламеллярные структуры, а образует массу мелких пузырьков, которые скапливаютсяя в отдельные зоны и даже могут формировать сложные решетчатые структуры (проламеллярные тела). В мембранах этиопластов содержится протохлорофилл, предшественник хлорофилла желтого цвета. Под действие света из этиопластов образуются хлоропласты, протохлорофилл превращается в хлорофилл, происходит синтез новых мембран, фотосинтетических ферментов и компонентов цепи переноса электронов.
При освещении клеток мембранные пузырьки и трубочки быстро реорганизуются, из них развивается полная система ламелл и тилакоидов, характерная для нормального хлоропласта.
Лейкопласты отличаются от хлоропластов отсутствием развитой ламеллярной системы (рис. 226 б). Встречаются они в клетках запасающих тканей. Из-за их неопределенной морфологии лейкопласты трудно отличить от пропластид, а иногда и от митохондрий. Они, как и пропластиды, бедны ламеллами, но тем не менее способны к образованию под влиянием света нормальных тилакоидных структур и к приобретению зеленой окраски. В темноте лейкопласты могут накапливать в проламеллярных телах различные запасные вещества, а в строме лейкопластов откладываются зерна вторичного крахмала. Если в хлоропластах происходит отложение так называемого транзиторного крахмала, который присутствует здесь лишь во время ассимиляции СО2, то в лейкопластах может происходить истинное запасание крахмала. В некоторых тканях (эндосперм злаков, корневища и клубни) накопление крахмала в лейкопластах приводит к образованию амилопластов, сплошь заполненных гранулами запасного крахмала, расположенных в строме пластиды (рис. 226в).
Другой формой пластид у высших растений является хромопласт, окрашивающийся обычно в желтый свет в результате накопления в нем каротиноидов (рис. 226г). Хромопласты образуются из хлоропластов и значительно реже их лейкопластов (например, в корне моркови). Процесс обесцвечивания и изменения хлоропластов легко наблюдать при развитии лепестков или при созревании плодов. При этом в пластидах могут накапливаться окрашенные в желтый цвет капельки (глобулы) или в них появляются тела в форме кристаллов. Эти процессы сопряжены с постепенным уменьшением числа мембран в пластиде, с исчезновением хлорофилла и крахмала. Процесс образования окрашенных глобул объясняется тем, что при разрушении ламелл хлоропластов выделяются липидные капли, в которых хорошо растворяются различные пигменты (например, каротиноиды). Таким образом, хромопласты представляют собой дегенерирующие формы пластид, подвернутые липофанерозу – распаду липопротедных комплексов.